Электрическая активность клеток миокарда
В естественных условиях клетки миокарда находятся в состоянии ритмической активности (возбуждения), поэтому об их потенциале покоя можно говорить лишь условно. У большинства клеток он составляет около 90 мВ и определяется почти целиком концентрационным градиентом ионов К+.
Потенциалы действия (ПД), зарегистрированные в разных отделах сердца при помощи внутриклеточных микроэлектродов, существенно различаются по форме, амплитуде и длительности (рис. 7.3, А). На рис. 7.3, Б схематически показан ПД одиночной клетки миокарда желудочка. Для возникновения этого потенциала потребовалось деполяризовать мембрану на 30 мВ. В ПД различают следующие фазы: быструю начальную деполяризацию — фаза 1; медленную реполяризацию, так называемое плато — фаза 2; быструю реполяризацию — фаза 3; фазу покоя — фаза 4.
Фаза 1 в клетках миокарда предсердий, сердечных проводящих миоцитов (волокна Пуркинье) и миокарда желудочков имеет ту же природу, что и восходящая фаза ПД нервных и скелетных мышечных волокон — она обусловлена повышением натриевой проницаемости, т. е. активацией быстрых натриевых каналов клеточной мембраны. Во время пика ПД происходит изменение знака мембранного потенциала (с —90 до +30 мВ).
Деполяризация мембраны вызывает активацию медленных натрий-кальциевых каналов. Поток ионов Са2+ внутрь клетки по этим каналам приводит к развитию плато ПД (фаза 2). В период плато натриевые каналы инактивируются и клетка переходит в состояние абсолютной рефрактерности. Одновременно происходит активация калиевых каналов. Выходящий из клетки поток ионов К+ обеспечивает быструю реполяризацию мембраны (фаза 3), во время которой кальциевые каналы закрываются, что ускоряет процесс реполяризации (поскольку падает входящий кальциевый ток, деполяризующий мембрану).
Реполяризация мембраны вызывает постепенное закрывание калиевых и реактивацию натриевых каналов. В результате возбудимость миокардиальной клетки восстанавливается — это период так называемой относительной рефрактерности.
В клетках рабочего миокарда (предсердия, желудочки) мембранный потенциал (в интервалах между следующими друг за другом ПД) поддерживается на более или менее постоянном уровне. Однако в клетках синусно-предсердного узла, выполняющего роль водителя ритма сердца, наблюдается спонтанная диастолическая деполяризация (фаза 4), при достижении критического уровня которой (примерно —50 мВ) возникает новый ПД (см. рис. 7.3, Б). На этом механизме основана авторитмическая активность указанных сердечных клеток. Биологическая активность этих клеток имеет и другие важные особенности: 1) малую крутизну подъема ПД; 2) медленную реполяризацию (фаза 2), плавно переходящую в фазу быстрой реполяризации (фаза 3), во время которой мембранный потенциал достигает уровня —60 мВ (вместо —90 мВ в рабочем миокарде), после чего вновь начинается фаза медленной диастолической деполяризации. Сходные черты имеет электрическая активность клеток предсердно-желудочкового узла, однако скорость спонтанной диастолической деполяризации у них значительно ниже, чем у клеток синусно-предсердного узла, соответственно ритм их потенциальной автоматической активности меньше.
Ионные механизмы генерации электрических потенциалов в клетках водителя ритма полностью не расшифрованы. Установлено, что в развитии медленной диастолической деполяризации и медленной восходящей фазы ПД клеток синусно-предсердного узла ведущую роль играют кальциевые каналы. Они проницаемы не только для ионов Са2+, но и для ионов Na+. Быстрые натриевые каналы не принимают участия в генерации ПД этих клеток.
Скорость развития медленной диастолической деполяризации регулируется автономной (вегетативной) нервной системой. В случае влияния симпатической части медиатор норадреналин активирует медленные кальциевые каналы, вследствие чего скорость диастолической деполяризации увеличивается и ритм спонтанной активности возрастает. В случае влияния парасимпатической части медиатор АХ повышает калиевую проницаемость мембраны, что замедляет развитие диастолической деполяризации или прекращает ее, а также гиперполяризует мембрану. По этой причине происходит урежение ритма или прекращение автоматии.
Способность клеток миокарда в течение жизни человека находиться в состоянии непрерывной ритмической активности обеспечивается эффективной работой ионных насосов этих клеток. В период диастолы из клетки выводятся ионы Na+, а в клетку возвращаются ионы К+. Ионы Са2+, проникшие в цитоплазму, поглощаются эндоплазматической сетью. Ухудшение кровоснабжения миокарда (ишемия) ведет к обеднению запасов АТФ и креатинфосфата в миокардиальных клетках; работа насосов нарушается, вследствие чего уменьшается электрическая и механическая активность миокардиальных клеток.
Функции проводящей системы сердца
Спонтанная генерация ритмических импульсов является результатом слаженной деятельности многих клеток синусно-предсердного узла, которая обеспечивается тесными контактами (нексусы) и электротоническим взаимодействием этих клеток. Возникнув в синусно-предсердном узле, возбуждение распространяется по проводящей системе на сократительный миокард.
Особенностью проводящей системы сердца является способность каждой клетки самостоятельно генерировать возбуждение. Существует так называемый градиент автоматии, выражающийся в убывающей способности к автоматии различных участков проводящей системы по мере их удаления от синусно-предсердного узла, генерирующего импульса с частотой до 60—80 в минуту.
В обычных условиях автоматия всех нижерасположенных участков проводящей системы подавляется более частыми импульсами, поступающими из синусно-предсердного узла. В случае поражения и выхода из строя этого узла водителем ритма может стать предсердно-желудочковый узел. Импульсы при этом будут возникать с частотой 40—50 в минуту. Если окажется выключенным и этот узел, водителем ритма могут стать волокна предсердно-желудочкового пучка (пучок Гиса). Частота сердечных сокращений в этом случае не превысит 30—40 в минуту. Если выйдут из строя и эти водители ритма, то процесс возбуждения спонтанно может возникнуть в клетках волокон Пуркинье. Ритм сердца при этом будет очень редким — примерно 20 в минуту.
Отличительной особенностью проводящей системы сердца является наличие в ее клетках большого количества межклеточных контактов — нексусов. Эти контакты являются местом перехода возбуждения с одной клетки на другую. Такие же контакты имеются и между клетками проводящей системы и рабочего миокарда. Благодаря наличию контактов миокард, состоящий из отдельных клеток, работает как единой целое. Существование большого количества межклеточных контактов увеличивает надежность проведения возбуждения в миокарде.
Возникнув в синусно-предсердном узле, возбуждение распространяется по предсердиям, достигая предсердно-желудочкового (атриовентрикулярного) узла. В сердце теплокровных животных существуют специальные проводящие пути между синусно-предсердным и предсердно-желудочковым узлами, а также между правым и левым предсердиями. Скорость распространения возбуждения в этих проводящих путях ненамного превосходит скорость распространения возбуждения по рабочему миокарду. В предсердно-желудочковом узле благодаря небольшой толщине его мышечных волокон и особому способу их соединения возникает некоторая задержка проведения возбуждения. Вследствие задержки возбуждение доходит до предсердно-желудочкового пучка и сердечных проводящих миоцитов (волокна Пуркинье) лишь после того, как мускулатура предсердий успевает сократиться и перекачать кровь из предсердий в желудочки.
Следовательно, атриовентрикулярная задержка обеспечивает необходимую последовательность (координацию) сокращений предсердий и желудочков.
Скорость распространения возбуждения в предсердно-желудочковом пучке и в диффузно расположенных сердечных проводящих миоцитах достигает 4,5—5 м/с, что в 5 раз больше скорости распространения возбуждения по рабочему миокарду. Благодаря этому клетки миокарда желудочков вовлекаются в сокращение почти одновременно, т. е. синхронно (см. рис. 7.2). Синхронность сокращения клеток повышает мощность миокарда и эффективность нагнетательной функции желудочков. Если бы возбуждение проводилось не через предсердно-желудочковый пучок, а по клеткам рабочего миокарда, т. е. диффузно, то период асинхронного сокращения продолжался бы значительно дольше, клетки миокарда вовлекались в сокращение не одновременно, а постепенно и желудочки потеряли бы до 50% своей мощности.
Таким образом, наличие проводящей системы обеспечивает ряд важных физиологических особенностей сердца: 1) ритмическую генерацию импульсов (потенциалов действия); 2) необходимую последовательность (координацию) сокращений предсердий и желудочков; 3) синхронное вовлечение в процесс сокращения клеток миокарда желудочков (что увеличивает эффективность систолы).
ФИЗИОЛОГИЯ СЕРДЦА
Самой главной функцией сердца является насосная. т. е. способность сердца непрерывно перекачивать кровь из вен в артерии, из большого круга кровообращения в малый. Цель этого насоса – доставлять кровь, несущую кислород и питательные вещества, ко всем органам и тканям, чтобы обеспечить их жизнедеятельность, забрать вредные продукты жизнедеятельности и донести их к обезвреживающим органам.
Сердце – это своеобразный вечный двигатель. В этом и последующих выпусках по физиологии сердца будут описаны сложнейшие механизмы, за счёт которых оно функционирует.
Выделяют 4 основные свойства сердечной ткани:
- Возбудимость – способность отвечать на действия раздражителей возбуждением в виде электрических импульсов.
- Автоматизм – способность самовозбуждаться, т. е. генерировать электрические импульсы в отсутствие внешних раздражителей.
- Проводимость – способность проводить возбуждение от клетки к клетке без затухания.
- Сократимость – способность мышечных волокон укорачиваться или увеличивать своё напряжение.
Средняя оболочка сердца – миокард – состоит из клеток, которые называются кардиомиоцитами. Кардиомиоциты не все одинаковы по своей структуре и выполняют различные функции. Выделяют следующие разновидности кардиомиоцитов:
- Сократительные (рабочие, типичные) кардиомиоциты составляют 99 % массы миокарда и обеспечивают непосредственно сократительную функцию сердца.
- Проводящие (атипичные, специализированные) кардиомиоциты. которые формируют проводящую систему сердца. Среди проводящих кардиомиоцитов различают 2 вида клеток – Р-клетки и клетки Пуркинье. Р-клетки (от англ. рale – бледный) обладают способностью периодически генерировать электрические импульсы, чем и обеспечивают функцию автоматизма. Клетки Пуркинье обеспечивают проведение импульсов ко всем отделам миокарда и имеют слабую способность к автоматизму.
- Переходные кардиомиоциты или Т-клетки (от англ. transitional — переходный) расположены между проводящими и сократительными кардиомиоцитами и обеспечивают их взаимодействие (т. е. передачу импульса от проводящих клеток к сократительным).
- Секреторные кардиомиоциты расположены преимущественно в предсердиях. Они выделяют в просвет предсердий натрийуретический пептид – гормон, регулирующий водно-электролитный баланс в организме и артериальное давление.
Все типы клеток миокарда не обладают способностью к делению, т. е. не способны к регенерации. Если у человека повышается нагрузка на сердце (например, у спортсменов), увеличение мышечной массы происходит за счёт увеличения объёма отдельных кардиомиоцитов (гипертрофии), а не их общего количества (гиперплазии).
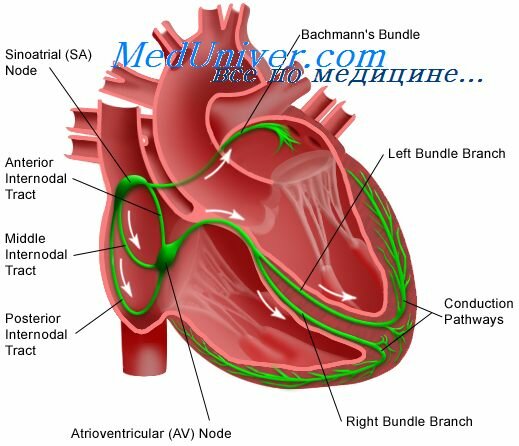
Теперь рассмотрим подробнее строение проводящей системы сердца (рис. 1). Она включает в себя следующие основные структуры:
- Синоатриальный (от латинского sinus – пазуха, atrium – предсердие), или синусовый , узел расположен на задней стенке правого предсердия около устья верхней полой вены. Он образован P-клетками, которые посредством Т-клеток связаны между собой и с сократительными кардиомиоцитами предсердий. От синоатриального узла в направлении к атриовентрикулярному узлу отходят 3 межузловых пучка: передний (пучок Бахмана), средний (пучок Венкебаха) и задний (пучок Тореля).
- Атриовентрикулярный (от лат. аtrium – предсердие, ventriculum – желудочек) узел – расположен в зоне перехода от предсердных кардиомиоцитов к пучку Гиса. Содержит Р-клетки, но в меньшем количестве, чем в синусовом узле, клетки Пуркинье, Т-клетки.
- Предсердно-желудочковый пучок, или пучок Гиса (описан немецким анатомом В. Гисом в 1893 г.) в норме является единственным путём проведения возбуждения от предсердий к желудочкам. Он отходит от атриовентрикулярного узла общим стволом и проникает в межжелудочковую перегородку. Здесь пучок Гиса делится на 2 ножки – правую и левую, идущие к соответствующим желудочкам. Левая ножка делится на 2 ветви – передневерхнюю и задненижнюю. Ветви пучка Гиса заканчиваются в желудочках сетью мелких волокон Пуркинье (описаны чешским физиологом Я. Пуркинье в 1845 г.).
1. Синусовый узел. 2. Атриовентрикулярный узел. 3. Ножки пучка Гиса. 4. Волокна Пуркинье.
У некоторых людей встречаются дополнительные (аномальные) проводящие пути (пучок Джеймса, пучок Кента), которые участвуют в возникновении нарушений сердечного ритма (например, синдрома преждевременного возбуждения желудочков).
В норме возбуждение зарождается в синусном узле, переходит на миокард предсердий, и, пройдя атриовентрикулярный узел, распространяется по ножкам пучка Гиса и волокнам Пуркинье на миокард желудочков.
Таким образом, нормальный ритм сердца определяется активностью синоатриального узла, который называют водителем ритма первого порядка, или истинным пейсмекером (от англ. pacemaker – «отбивающий шаг»). Автоматизм присущ также другим структурам проводящей системы сердца. Водитель второго порядка локализован в атриовентрикулярном узле. Водителями третьего порядка являются клетки Пуркинье, входящие в состав проводящей системы желудочков.
Продолжение следует.
В рассылке использованы материалы учебного пособия «Физиология сердца» под ред. акад. Б. И. Ткаченко.
Проводящая система сердца. Синусовый узел
На рисунке показана схема проводящей системы сердца. В ее состав входят: (1) синусный узел (который также называют синоатриальным или С-А узлом), где и происходит ритмическая генерация импульсов; (2) предсердные межузловые пучки, по которым импульсы проводятся от синусного узла к агриовентрикулярному узлу; (3) атриовентрикулярный узел, в котором происходит задержка проведения импульсов от предсердий к желудочкам; (4) атриовентрикулярный пучок, по которому импульсы проводятся к желудочкам; (5) левая и правая ножки А-В пучка, состоящие из волокон Пуркинье, благодаря которым импульсы достигают сократительного миокарда.
Синусный (синоатриальный) узел представляет собой небольшую эллипсовидную пластинку шириной 3 мм, длиной 15 мм и толщиной 1 мм, состоящую из атипических кардиомноцитов. С-А узел расположен в верхней части заднебоковой стенки правого предсердия у места впадения в него верхней полой вены. Клетки, входящие в состав С-А узла, практически не содержат сократительных филаментов; их диаметр всего лишь 3-5 мкм (в отличие от предсердных сократительных волокон, диаметр которых 10-15 мкм). Клетки синусного узла непосредственно связаны с сократительными мышечными волокнами, поэтому потенциал действия, возникший в синусном узле, немедленно распространяется на миокард предсердий.
Автоматия — это способность некоторых сердечных волокон самостоятельно возбуждаться и вызывать ритмические сокращения сердца. Способностью к автоматии обладают клетки проводящей системы сердца, в том числе клетки синусного узла. Именно С-А узел контролирует ритм сердечных сокращений, как мы увидим далее. А сейчас обсудим механизм автоматии.
Механизм автоматии синусного узла. На рисунке представлены потенциалы действия клетки синусного узла, записанные на протяжении трех сердечных циклов, и для сравнения — одиночный потенциал действия кардиомиоцита желудочка. Необходимо отметить, что потенциал покоя клетки синусного узла имеет меньшую величину (от -55 до -60 мВ) в отличие от типичного кардиомиоцита (от -85 до -90 мВ). Это различие объясняется тем, что мембрана узловой клетки в большей степени проницаема для ионов натрия и кальция. Вход этих катионов в клетку нейтрализует часть внутриклеточных отрицательных зарядов и уменьшает величину потенциала покоя.
Прежде чем перейти к механизму автоматии. необходимо вспомнить, что в мембране кардиомиоцитов существуют три типа ионных каналов, которые играют важную роль в генерации потенциала действия: (1) быстрые натриевые каналы, (2) медленные Na+/Са2+-каналы, (3) калиевые каналы. В клетках миокарда желудочков кратковременное открытие быстрых натриевых каналов (на несколько десятитысячных долей секунды) и вход ионов натрия в клетку приводит к быстрой деполяризации и перезарядке мембраны кардиомиоцита. Фаза плато потенциала действия, которая продолжается 0,3 сек, формируется за счет открытия медленных Na+/Ca -каналов. Затем открываются калиевые каналы, происходит диффузия ионов калия из клетки — и мембранный потенциал возвращается к исходному уровню.
В клетках синусного узла потенциал покоя меньше, чем в клетках сократительного миокарда (-55 мВ вместо -90 мВ). В этих условиях ионные каналы функционируют по-другому. Быстрые натриевые каналы инактивированы и не могут участвовать в генерации импульса. Дело в том, что любое уменьшение мембранного потенциала до -55 мВ на срок больший, чем несколько миллисекунд, приводит к закрытию инактивационных ворот во внутренней части быстрых натриевых каналов. Большая часть этих каналов оказывается полностью блокирована. В этих условиях могут открыться только медленные Na+/Ca -каналы, и поэтому именно их активация становится причиной возникновения потенциала действия. Кроме того, активация медленных Na/Ca -каналов обусловливает сравнительно медленное развитие процессов деполяризации и реполяризации в клетках синусного узла в отличие от волокон сократительного миокарда желудочков.
Оглавление темы «Проводящая система сердца. ЭКГ»:




